中国畜禽种业 ›› 2026, Vol. 22 ›› Issue (5): 52-60.doi: 10.19543/j.cnki.1673-4556.20260329.003
周宁1,2,3( ), 刘锦玉2, 卢立志2, 曾涛2, 郑嫩珠1, 张钰3, 李丽1(), 顾天天1,2()
), 刘锦玉2, 卢立志2, 曾涛2, 郑嫩珠1, 张钰3, 李丽1(), 顾天天1,2()
Ning Zhou1,2,3(), Jinyu Liu2, Lizhi Lu2, Tao Zeng2, Nenzhu Zheng1, Yu Zhang3, Li Li1(), Tiantian Gu1,2()
摘要:
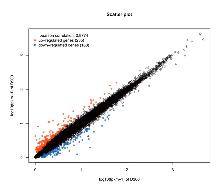
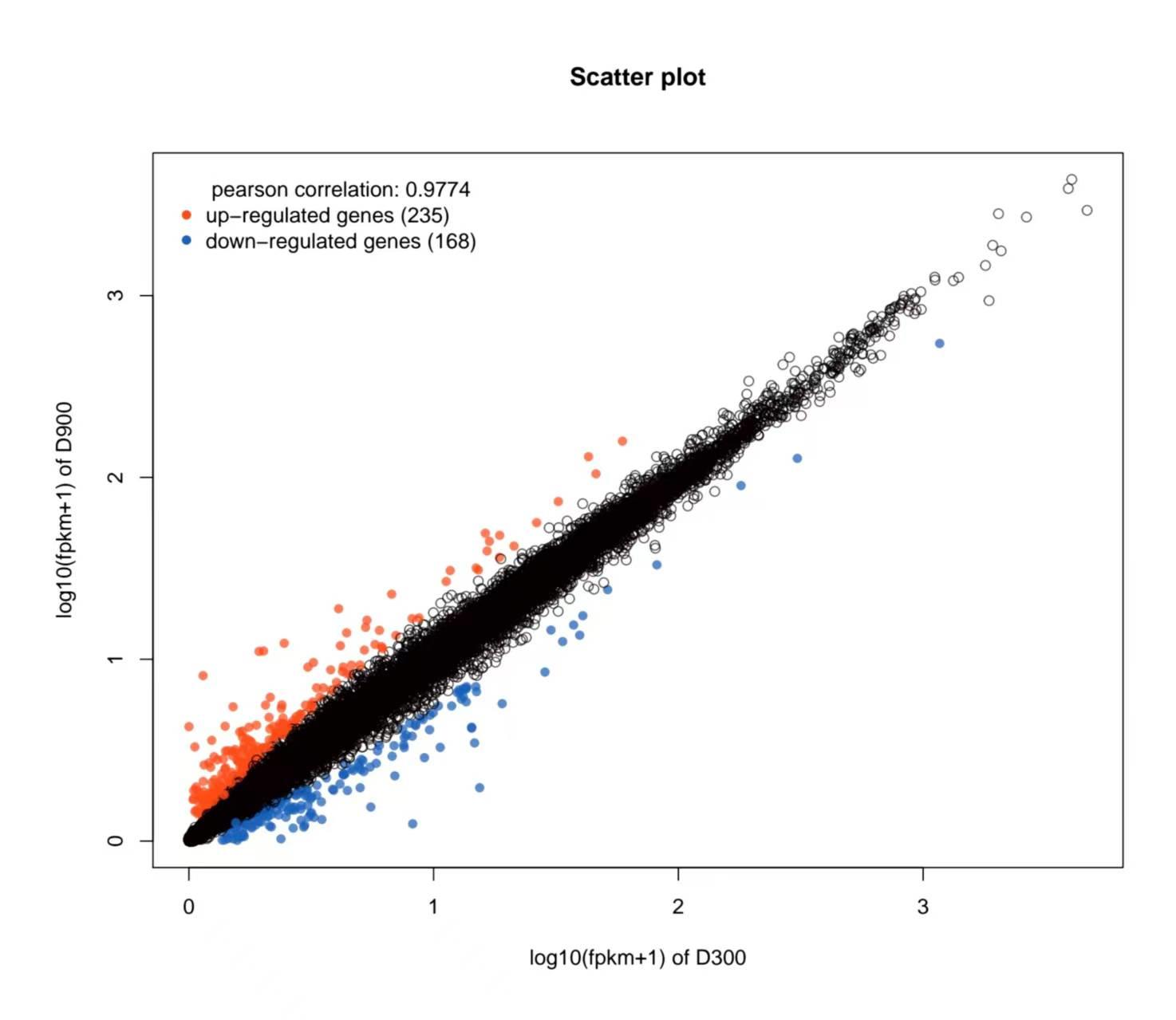
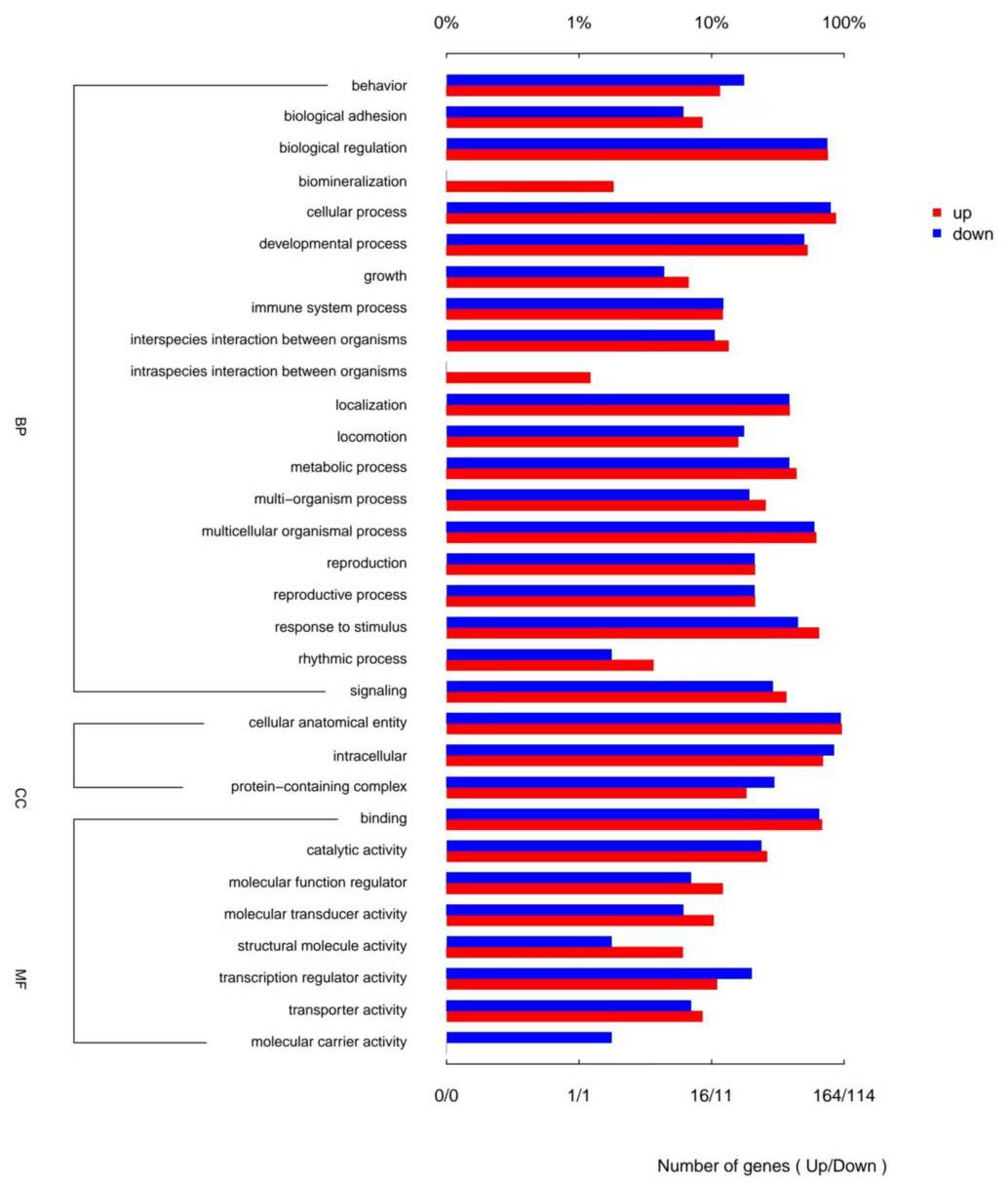
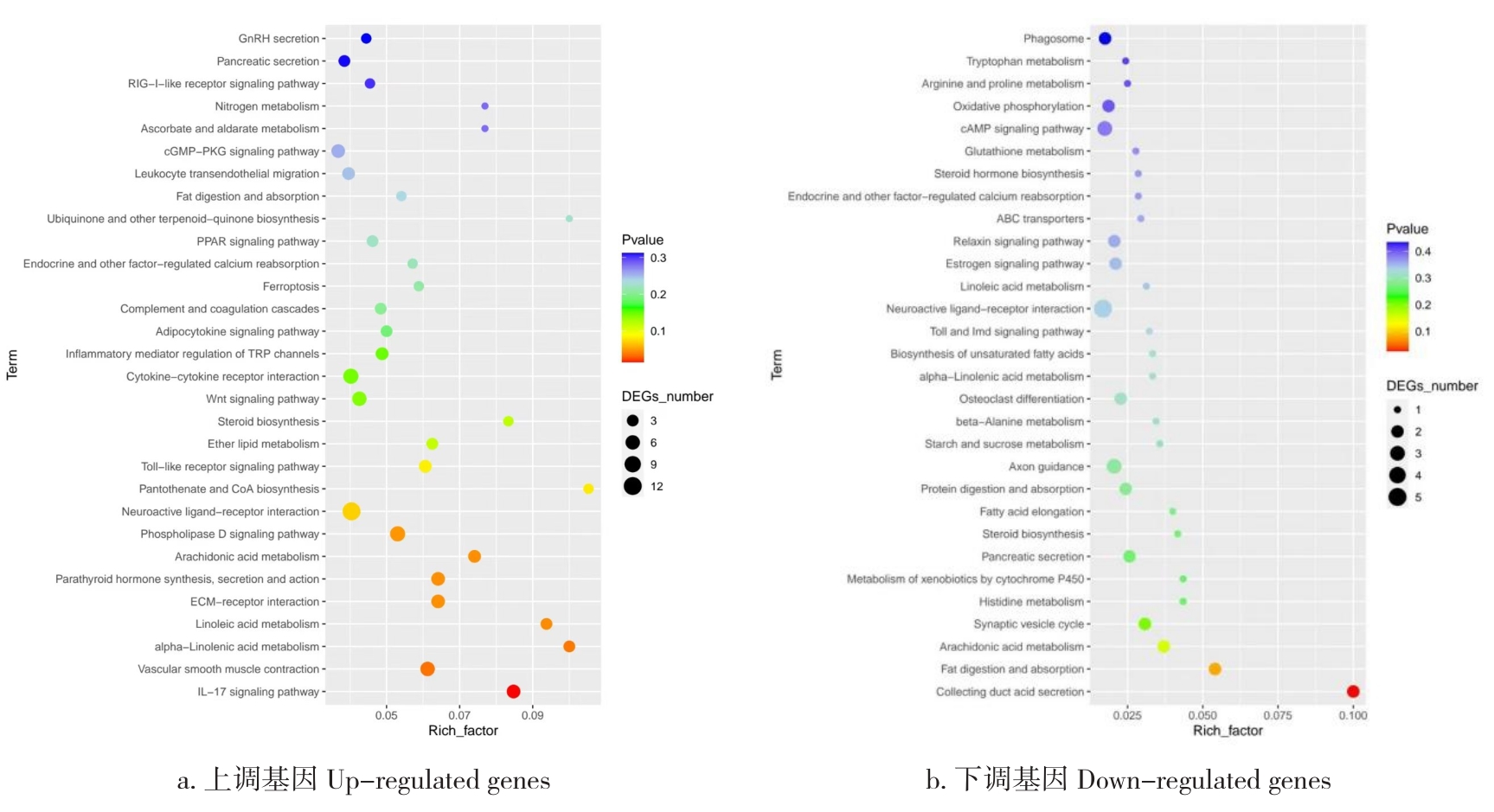
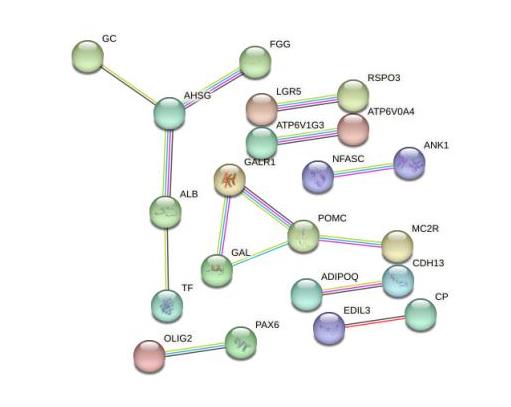
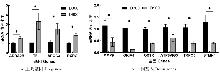
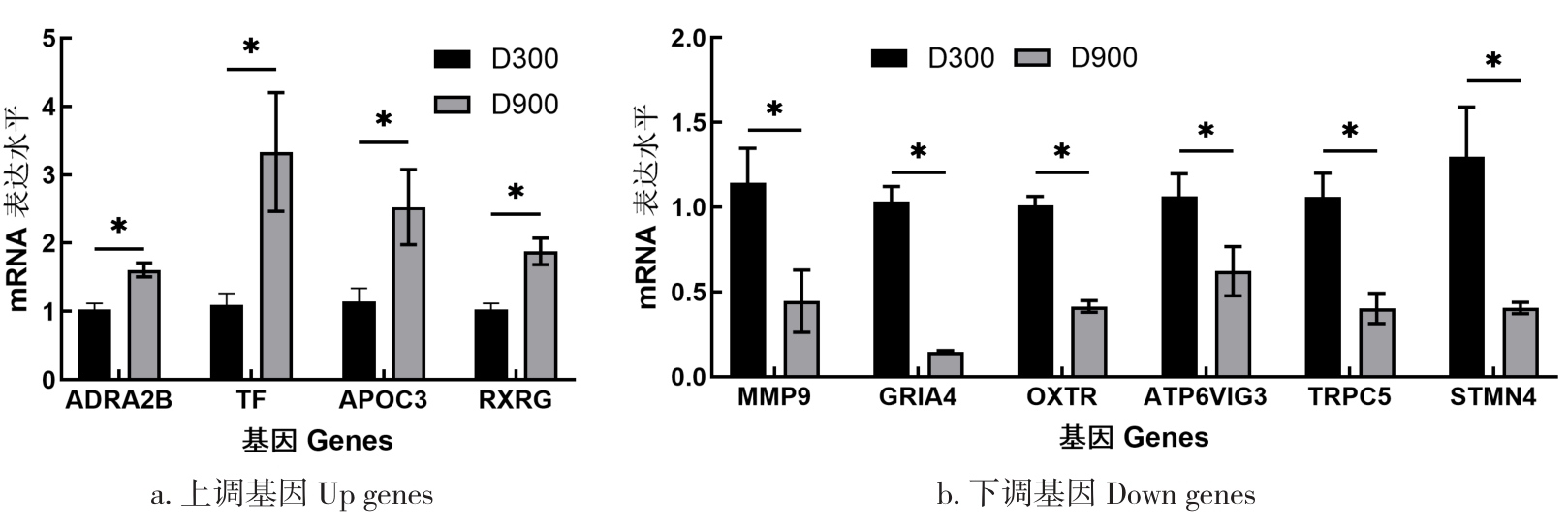
目的 该试验旨在研究300日龄和900日龄蛋鸭卵巢组织转录本的差异,并筛选影响卵巢衰老的关键基因或信号通路。 方法 该试验采集300日龄青年蛋鸭和900日龄老年蛋鸭卵巢组织,通过Illumina HiSeq平台进行深度测序,并对差异表达基因(Differentially expressed genes, DEGs)进行基因本体(Gene ontology, GO)功能注释、京都基因与基因组百科全书(Kyoto encyclopedia of genes and genomes, KEGG)富集分析和实时荧光定量PCR(Quantitative Real-time PCR)验证。 结果 结果显示,900日龄和300日龄蛋鸭卵巢间共鉴定出403个基因表达存在显著差异,其中235个上调基因,168个下调基因。GO功能显著富集在代谢过程、免疫系统过程、生殖过程和转录调节活性等条目;KEGG通路显著富集在白细胞介素-17信号通路、ECM受体相互作用和花生四烯酸代谢等通路。qRT-PCR结果发现,10个基因(ADRA2B、TF、APOC3、RXRG、MMP9、GRIA4、OXTR、ATP6V1G3、TRPC5和STMN4)在900日龄和300日龄蛋鸭卵巢组织间表达变化模式与转录组测序结果一致,表明RNA-Seq结果可靠。 结论 转录组测序筛选出403个显著差异基因,其中TF、APOC3、TRPC5等基因可能与卵巢衰老相关。KEGG富集显示的白介素-17信号通路、ECM受体相互作用和花生四烯酸代谢等通路,可影响炎症反应和损伤,参与蛋鸭卵巢衰老。
中图分类号:
| [1] | 孙泽才, 陈新华, 黄凤娇, 等. 我国水禽产品市场基本特征分析[J]. 南方农业, 2015, 9(9): 103-105. |
| SUN Z C, CHEN X H, HUANG F J, et al. Analysis on the basic characteristics of waterfowl products market in China[J]. South China Agriculture, 2015, 9(9): 103-105. | |
| [2] | 冯佩诗. 全基因组重测序及转录组测序揭示鸭种群结构及驯化适应的分子基础[D]. 广州: 华南农业大学, 2018. |
| FENG P S. Whole-genome resequencing and transcriptome sequencing reveal population structure and domestication signature of ducks[D]. Guangzhou: South China Agricultural University, 2018. | |
| [3] | ALIMADADI A, MUNROE P B, JOE B, et al. Meta-analysis of dilated cardiomyopathy using cardiac RNA-seq transcriptomic datasets[J]. Genes, 2020, 11(1): 60. |
| [4] | 王珍珍, 芦治学, 屈元启, 等. 不同品种蛋鸭蛋品质比较分析[J]. 中国家禽, 2019, 41(12): 55-58. |
| WANG Z Z, LU Z X, QU Y Q, et al. Analysis on egg quality traits of four laying duck breeds[J]. China Poultry, 2019, 41(12): 55-58. | |
| [5] | ZHOU J, YU J Z, ZHU M Y, et al. Genome-wide association analysis and genetic parameters for egg production traits in Peking ducks[J]. Animals, 2024, 14(13): 1891. |
| [6] | CHANG Y G, GUO R B, ZENG T, et al. Analysis of transcriptomic differences in the ovaries of high- and low-laying ducks[J]. Genes, 2024, 15(2): 181. |
| [7] | WANG S, ZHENG Y X, LI J Y, et al. Single-cell transcriptomic atlas of primate ovarian aging[J]. Cell, 2020, 180(3): 585-600.e19. |
| [8] | RUSS J E, HAYWOOD M E, LANE S L, et al. Spatially resolved transcriptomic profiling of ovarian aging in mice[J]. iScience, 2022, 25(8): 104819. |
| [9] | ZHANG J J, CHEN Q, DU D F, et al. Can ovarian aging be delayed by pharmacological strategies[J]. Aging, 2019, 11(2): 817-832. |
| [10] | ISOLA J V V, OCAÑAS S R, HUBBART C R, et al. A single-cell atlas of the aging mouse ovary[J]. Nature Aging, 2024, 4(1): 145-162. |
| [11] | MAY-PANLOUP P, BOUCRET L, CHAO DE LA BARCA J M, et al. Ovarian ageing: The role of mitochondria in oocytes and follicles[J]. Human Reproduction Update, 2016, 22(6): 725-743. |
| [12] | LIM J, LUDERER U. Oxidative damage increases and antioxidant gene expression decreases with aging in the mouse ovary[J]. Biology of Reproduction, 2011, 84(4): 775-782. |
| [13] | ZHU J, YANG Q L, LI H, et al. Sirt3 deficiency accelerates ovarian senescence without affecting spermatogenesis in aging mice[J]. Free Radical Biology & Medicine, 2022, 193(Pt 2): 511-525. |
| [14] | GONG H X, QIU X T, LI P P, et al. Immune infiltration analysis reveals immune cell signatures in salivary gland tissue of primary Sjögren’s syndrome[J]. Frontiers in Medicine, 2023, 10: 1033232. |
| [15] | 陈黎, 李国勤, 田勇, 等. 北京鸭腹部脂肪组织的转录组特征分析[J]. 浙江农业学报, 2016, 28(5): 743-747. |
| CHEN L, LI G Q, TIAN Y, et al. Transcriptome analysis of abdominal fats from Peking ducks by RNA-seq[J]. Acta Agriculturae Zhejiangensis, 2016, 28(5): 743-747. | |
| [16] | GRABHERR M G, HAAS B J, YASSOUR M, et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome[J]. Nature Biotechnology, 2011, 29(7): 644-652. |
| [17] | LAN D L, XIONG X R, WEI Y L, et al. RNA-Seq analysis of yak ovary: Improving yak gene structure information and mining reproduction-related genes[J]. Science China Life Sciences, 2014, 57(9): 925-935. |
| [18] | WESTERMANN A J, VOGEL J. Cross-species RNA-seq for deciphering host–microbe interactions[J]. Nature Reviews Genetics, 2021, 22(6): 361-378. |
| [19] | JIN C L, WANG S L, WANG S, et al. Age-related calcium signaling disturbance restricted cAMP metabolism and induced ovarian oxidation stress in laying ducks[J]. Poultry Science, 2025, 104(1): 104551. |
| [20] | REN J D, SUN C S, CHEN L, et al. Exploring differentially expressed key genes related to development of follicle by RNA-seq in Peking ducks (Anas Platyrhynchos)[J]. PLoS One, 2019, 14(6): e0209061. |
| [21] | TAO W J, JIN F, FAN Q W, et al. Effects of chitosan oligosaccharide on production performance, egg quality and ovarian function in laying hens with fatty liver syndrome[J]. Animals, 2022, 12(18): 2465. |
| [22] | 李娟, 白雪, 杨梦丽, 等. 单细胞转录组测序技术在反刍动物卵巢生长发育中的调控研究[J]. 中国畜禽种业, 2025, 21(9): 22-28. |
| LI J, BAI X, YANG M L, et al. Single-cell transcriptome sequencing technology in the regulatory research of ovarian growth and development in ruminant animals[J]. The Chinese Livestock and Poultry Breeding, 2025, 21(9): 22-28. | |
| [23] | ZENG Y T, WANG C, YANG C T, et al. Unveiling the role of chronic inflammation in ovarian aging: insights into mechanisms and clinical implications[J]. Human Reproduction, 2024, 39(8): 1599-1607. |
| [24] | LI Y, CHANG L H, HUANG W Q, et al. IL-17A mediates pyroptosis via the ERK pathway and contributes to steroid resistance in CRSwNP[J]. Journal of Allergy and Clinical Immunology, 2022, 150(2): 337-351. |
| [25] | BOROWCZYK J, SHUTOVA M, BREMBILLA N C, et al. IL-25 (IL-17E) in epithelial immunology and pathophysiology[J]. The Journal of Allergy and Clinical Immunology, 2021, 148(1): 40-52. |
| [26] | MIOSSEC P, KOLLS J K. Targeting IL-17 and TH17 cells in chronic inflammation[J]. Nature Reviews Drug Discovery, 2012, 11(10): 763-776. |
| [27] | KIM S K, JUNG S M, PARK K S, et al. Integrative analysis of lung molecular signatures reveals key drivers of idiopathic pulmonary fibrosis[J]. BMC Pulmonary Medicine, 2021, 21(1): 404. |
| [28] | JIAO X Y, JIAO Y X, CUI J W, et al. S100A4 targets PPP1CA/IL-17 to inhibit the senescence of sheep endometrial epithelial cells[J]. Frontiers in Veterinary Science, 2024, 11: 1466482. |
| [29] | WANG H, ZHU X Q, SHEN J Y, et al. Quantitative iTRAQ-based proteomic analysis of differentially expressed proteins in aging in humans and monkey[J]. BMC Genomics, 2019, 20(1): 725. |
| [30] | SUTHERLAND T E, DYER D P, ALLEN J E. The extracellular matrix and the immune system: A mutually dependent relationship[J]. Science, 2023, 379(6633): eabp8964. |
| [31] | HUA X M, YU L F, ZHU H, et al. Research progress of circRNAs in bone-related diseases[J]. Frontiers in Oncology, 2025, 15: 1481322. |
| [32] | EDWARDS J M, MCCARTHY C G, WENCESLAU C F. The obligatory role of the acetylcholine-induced endothelium-dependent contraction in hypertension: Can arachidonic acid resolve this inflammation[J]. Current Pharmaceutical Design, 2020, 26(30): 3723-3732. |
| [33] | ZHAO H K, WU L, YAN G F, et al. Inflammation and tumor progression: Signaling pathways and targeted intervention[J]. Signal Transduction and Targeted Therapy, 2021, 6: 263. |
| [34] | BAKHTIARI S, AHMADI B, ASRI N, et al. Unraveling the serum protein landscape in celiac disease: current evidence and future directions[J]. Immunity, Inflammation and Disease, 2025, 13(5): e70169. |
| [35] | LEE A R, PECHENINO A S, DONG H, et al. Aging, estrogen loss and epoxyeicosatrienoic acids (EETs)[J]. PLoS One, 2013, 8(8): e70719. |
| [36] | YAO Y, WANG B, JIANG Y B, et al. The mechanisms crosstalk and therapeutic opportunities between ferroptosis and ovary diseases[J]. Frontiers in Endocrinology, 2023, 14: 1194089. |
| [37] | SZE S C W, ZHANG L, ZHANG S Q, et al. Aberrant transferrin and ferritin upregulation elicits iron accumulation and oxidative inflammaging causing ferroptosis and undermines estradiol biosynthesis in aging rat ovaries by upregulating NF-κb-activated inducible nitric oxide synthase: first demonstration of an intricate mechanism[J]. International Journal of Molecular Sciences, 2022, 23(20): 12689. |
| [38] | HASHEMITABAR M, BAHMANZADEH M, MOSTAFAIE A, et al. A proteomic analysis of human follicular fluid: comparison between younger and older women with normal FSH levels[J]. International Journal of Molecular Sciences, 2014, 15(10): 17518-17540. |
| [39] | PRAKASH T P, MULLICK A E, RINEY S, et al. Discovery of long-acting APOC3 siRNA for treating patients with hypertriglyceridemia[J]. Nucleic Acids Research, 2025, 53(20): gkaf1063. |
| [40] | PACKARD C J, TASKINEN M R, BJÖRNSON E, et al. Genetically determined increase in apolipoprotein C-III (APOC3 gain-of-function) delays very low-density lipoprotein clearance in humans[J]. Atherosclerosis, 2025, 404: 119166. |
| [41] | ZEWINGER S, REISER J, JANKOWSKI V, et al. Apol-ipoprotein C3 induces inflammation and organ damage by alternative inflammasome activation[J]. Nature Immunology, 2020, 21(1): 30-41. |
| [42] | NAVARRO-PANDO J M, ALCOCER-GÓMEZ E, CASTEJÓN-VEGA B, et al. Inhibition of the NLRP3 inflammasome prevents ovarian aging[J]. Science Advances, 2021, 7(1): eabc7409. |
| [43] | MARTÍN-GONZÁLEZ C, MARTÍN-FOLGUERAS T, QUE-VEDO-ABELEDO J C, et al. Disease activity in patients with rheumatoid arthritis increases serum levels of apolipoprotein C-III[J]. Clinical and Experimental Rheumatology, 2023, 41(1): 67-73. |
| [44] | BORNFELDT K E. Apolipoprotein C3: form begets function[J]. Journal of Lipid Research, 2024, 65(1): 100475. |
| [45] | ZHANG X Y, ZHANG L, XIANG W P. The impact of mitochon-drial dysfunction on ovarian aging[J]. Journal of Translational Medicine, 2025, 23(1): 211. |
| [46] | DEMAREE I S, KUMAR S, TENNESSEN K, et al. Effects of TRPC1's lysines on heteromeric TRPC5-TRPC1 channel function[J]. Cells, 2024, 13(23): 2019. |
| [47] | KECK M, HERMANN C, LÜTZEL K, et al. Photoswitchable TRPC6 channel activators evoke distinct channel kinetics reflecting different gating behaviors[J]. iScience, 2024, 27(10): 111008. |
| [48] | NAYLOR J, AL-SHAWAF E, MCKEOWN L, et al. TRPC5 channel sensitivities to antioxidants and hydroxylated stilbenes[J]. Journal of Biological Chemistry, 2011, 286(7): 5078-5086. |
| [49] | LI Z T, GUO G, WANG H R, et al. TRPC5 channel modulates endothelial cells senescence[J]. European Journal of Phar-macology, 2017, 802: 27-35. |
| [50] | ISOLA J V V, BISWAS S, JAYARATHNE H, et al. Cana-gliflozin treatment prevents follicular exhaustion and attenuates hallmarks of ovarian aging in genetically heterogenous mice[J]. GeroScience, 2025, 47(3): 3061-3076. |
| [51] | LIU X Y, ZHAO Y Q, FENG Y Z, et al. Ovarian aging: the silent catalyst of age-related disorders in female body[J]. Aging and Disease, 2025, 17(1): 132-158. |
| [1] | 王红梅. 断奶日龄对陕北绒山羊公羔生长和屠宰性能的影响[J]. 中国畜禽种业, 2023, 19(2): 83-86. |
| [2] | 林映升. 养殖密度对育肥大白猪体重校正日龄、体尺与背膘厚的影响[J]. 中国畜禽种业, 2022, 18(1): 59-61. |
| [3] | 张汝. 全舍饲旱养蛋鸭综合技术措施分析[J]. 中国畜禽种业, 2021, 17(11): 164-165. |
|
||